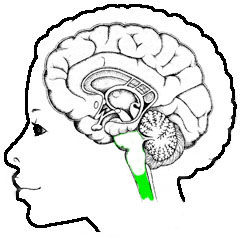
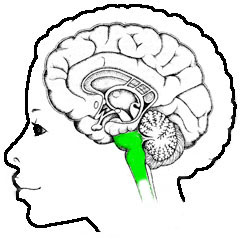
Neurologie, overzicht, hersenstam
Inleiding "Hersenstam" is de gebruikelijke verzamelnaam voor de meest basale elementen van
het brein
 , soms inclusief het ruggemerg en alles wat in het directe
verlengde daarvan ligt - maar meestal duidt "hersenstam" alleen op de onderdelen tussen
de emotionele hersenen en het ruggemerg. Ruggemerg en hersenstam vervullen de
meest basale regelfuncties van het lichaam, zoals blijkt uit het feit dat schade
eraan meestal ernstig, zeer moeilijk of niet te herstellen, en wat betreft de
hersenstam, snel dodelijk is. , soms inclusief het ruggemerg en alles wat in het directe
verlengde daarvan ligt - maar meestal duidt "hersenstam" alleen op de onderdelen tussen
de emotionele hersenen en het ruggemerg. Ruggemerg en hersenstam vervullen de
meest basale regelfuncties van het lichaam, zoals blijkt uit het feit dat schade
eraan meestal ernstig, zeer moeilijk of niet te herstellen, en wat betreft de
hersenstam, snel dodelijk is.
De gebruikelijke beschrijvingen van de hersenstam beginnen met de voor de
denkende mens meest belangrijke onderdelen, die aan de bovenkant ervan liggen. De beschrijving hier volgt
de algemene visie van deze website om zaken vanuit de evolutionaire
 kant te
bekijken. En in dit geval aangevuld met de wens om te laten zien hoe ruggemerg
en hersenstam werken in dienst van de rest van hersenen en het lichaam. Wat automatisch
betekent dat je van onderop moet
beginnen. Een noodzaak die bevestigd werd door de praktijk van eerdere pogingen
met de top-down benadering, die te omslachtig werden bij de steeds verdere
toevoeging van details aangaande uitvoering. kant te
bekijken. En in dit geval aangevuld met de wens om te laten zien hoe ruggemerg
en hersenstam werken in dienst van de rest van hersenen en het lichaam. Wat automatisch
betekent dat je van onderop moet
beginnen. Een noodzaak die bevestigd werd door de praktijk van eerdere pogingen
met de top-down benadering, die te omslachtig werden bij de steeds verdere
toevoeging van details aangaande uitvoering.
Geprobeerd is enige afwisseling aan
te brengen in technische details versus het waarom het zo in elkaar zit,
uitgaande van evolutionaire argumenten en wat praktisch plausibel lijkt.
Natuurlijk kan een enkele webpagina alleen de hoofdlijnen
geven, maar gepoogd is dusdanige informatie te verstrekken dat zelfstandig
verder zoeken in wat toch wel enigszins een doolhof is, met name aangaande
naamgeving waarvan wel drie of vier methoden zijn, makkelijker wordt gemaakt. Voor een overzicht
van die naamgeving, zie hier
 .
.
De opzet van deze pagina is om zelfstandig leesbaar te zijn,
maar een voorafgaande blik op Neurologie, overzicht, globaal
voor de algemene
structuur of Neurologie, neuronen, algemeen
kan behulpzaam
zijn. Neurologische (en andere) begrippen die elders kort toegelicht worden,
zijn vetgedrukt. De meeste van de bronverwijzingen (aangegeven met
) omtrent structuren zijn naar Wikipedia.
Opbouw
Het ruggemerg zelf is een product van voorgaande evolutie die begonnen is
met het ontstaan van verbindingscellen tussen
waarnemingsorganen en ledematen, gespecialiseerd tot neuronen
.
Naarmate gedurende de evolutie het bewegingstelsel ingewikkelder werd, met
ledematen met meerdere geledingen, werd ook het ruggemerg gecompliceerder, met
concentraties van neuronen voor de onderlinge coördinatie, en, heel belangrijk,
meer nauwkeurige besturing. Besturing wordt geregeld via het proces van
terugkoppeling
, dat wil zeggen: er zijn ook neuronen in de spieren van de ledematen die
krachten en posities terugmelden - in de neurologie heet dit de "proprioscopische"
informatie. De signalen
aan de spieren worden bijgesteld aan de hand van de signalen omtrent hoever het
ledemaat al gevorderd is in zijn geplande richting. En bij meerdere ledematen, denk
aan duizendpoten, moeten ook de ledematen onderling gecoördineerd worden. Ook
dit wordt door verzamelingen neuronen in het ruggemerg geregeld. De grotere
daarvan, die de ingewikkelde functies coördineren, zijn min of meer bolvormig en
heten ganglia.
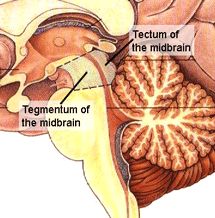
De grens tussen ruggemerg en hersenstam is enigszins vaag - waar het ruggemerg voornamelijk bestaat uit lange draden van uiteinden van neuronen (de axonen), met daartussen
gebieden met een aantal neuronknopen, wordt de hersenstam gedomineerd
door meer samenhangende structuren aangeduid als kernen, met veel
bedrading ertussen. In het eerste en tweede plaatje is de grens daarom wat vaag
is gehouden:
In de neurologie is het functioneren van het ruggemerg
veruit het meest bekende terrein, met bijvoorbeeld twaalf genummerde
hoofdzenuwbundels die alle signalen van en naar de rest van het lichaam
versturen, met per zenuwbundel bekend wat ze doen. De bijbehorende onderdelen
van het ruggemerg verzorgen de basale, zeg maar: mechanische, functies van het
lichaam, met als meest basaal het
bewegingsapparaat, en de bijbehorende reguleringsfuncties: ademhaling, hartslag,
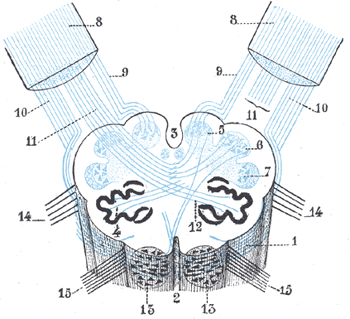
enzovoort. Zie onder voor een overzicht, een illustratie van de site van Ben
Best (voor een grotere versie zie aldaar
):
Goed zichtbaar zelfs in deze verkleinde vorm is dat het
ruggemerg ook de basale regulering van alle andere belangrijke
organen verzorgt. De onderdelen van het ruggemerg die dat doen vallen onder de term ganglia, oftewel
(zenuw)knopen, de kleinere structuren opgebouwd uit neuronen. Dit deel van het zenuwstelsel
heet het (ortho-)sympathische deel staande voor de activerende functies,
links in de afbeelding. Daarnaast is er het para-sympathische deel, rechts, aangestuurd door kernen in
de hersenstam, voor het reguleren van de functies in ruste, zoals de spijsvertering.
Van de twaalf zenuwbundels of
cranial nerves
van en naar de organen zijn in dit plaatje zichtbaar de oogzenuw (oculomotor nerve,
III), de gezichtzenuw (facial nerve, VII) en de zwervende zenuw (vagus,
X - naar diverse onderdelen in de borst).
Dit deel van het zenuwstelsel vervult zijn functie los van de hogere,
bewuste, delen van de hersenen, en heet daarom het autonome zenuwstelsel.
Aannemende dat dit allemaal goed werkt, heb je dus een goedwerkend
bewegingsapparaat. Wat beperkt bruikbaar als je niet weet waar je naar toe gaat.
Daarvoor is dus een waarnemingsapparaat nodig. Maar het heeft geen zin een
bewegings- plus waarnemingsapparaat te ontwikkelen als je geen idee hebt van wat je
er mee wilt doen. Waar je heen wilt. Er moet van begin af aan een ratio, een
overlevingswinst, gezeten hebben in welke beweging en waarneming dan ook, en hoe
primitief die ook mocht zijn.
In deze termen geformuleerd staat het antwoord er bijna al, in de term
"primitief": de winst bestond van
het begin af aan uit het wegwezen van een plaats waar gevaar dreigt.
 Wat
wel de vragen oproept wat "gevaar" is en hoe het primitieve leven dat ontdekt.
Het antwoord op die vragen is bekend uit een vroegere context dan die van het
zenuwstelsel: "gevaar" is het ontstaan van dode soortgenoten,
en de methode om dat te ontdekken is de afvalproducten van die dood. In de
primitieve wereld is het belangrijkste gevaar dat van "opgegeten worden",
en dat gaat in de meest primitieve vorm van "hap voor hap". Dat brengt
noodzakelijkerwijs restanten van het opgegeten wezen in de omgeving. Die
afvalproducten kunnen soortgenoten detecteren - dat is een biochemisch proces, wat nu heet "geur"
. (Overigens
verklaart dit ook waarom sommige roofvissen hun prooi in één keer inslikken, wat
qua digestie niet gunstig is - dit is ter vermijding van detectie). Wat
wel de vragen oproept wat "gevaar" is en hoe het primitieve leven dat ontdekt.
Het antwoord op die vragen is bekend uit een vroegere context dan die van het
zenuwstelsel: "gevaar" is het ontstaan van dode soortgenoten,
en de methode om dat te ontdekken is de afvalproducten van die dood. In de
primitieve wereld is het belangrijkste gevaar dat van "opgegeten worden",
en dat gaat in de meest primitieve vorm van "hap voor hap". Dat brengt
noodzakelijkerwijs restanten van het opgegeten wezen in de omgeving. Die
afvalproducten kunnen soortgenoten detecteren - dat is een biochemisch proces, wat nu heet "geur"
. (Overigens
verklaart dit ook waarom sommige roofvissen hun prooi in één keer inslikken, wat
qua digestie niet gunstig is - dit is ter vermijding van detectie).
Het ontdekken van afvalproducten van je eigen soort is een uitstekend signaal om naar elders te gaan, als je een
bewegingsapparaat hebt. Vandaar dat er ooit een bewegingsapparaat is ontstaan.
Al bij eencelligen, zie de afbeelding rechtboven. Het simpele motto was: "Wegwezen". Bestemming
onbelangrijk.
Andere primitieve omstandigheden verbonden aan "Gevaar!" zijn die van
temperatuur en tast. Werd je betast: wegwezen. En hitte is altijd een groot
gevaar geweest voor welk leven wezen dan ook. De waarneming daarvan heet het somatosensory system
(dus van de huid
e.d.). Het signaal dat dit systeem afgeeft, noemt de mens "pijn". In
de neurologie heet dit "nociceptie"
- in de huid zitten nociceptoren, die
mechanische, thermische of chemische schade detecteren en doorsturen naar het
ruggemerg. Aldaar worden acties ondernomen en signalen verder gestuurd, en die
in het bewustzijn vertaald zijn als de ervaring van "pijn".
Ook duidelijk evolutionair voordelig is het als je weet waar je heen moet, al
was het maar om zo niet juist in de richting van het roofdier te gaan. Waarvoor
een beeld van de omgeving gewenst is. Met allerlei tussenstappen die begonnen
met het kunnen detecteren van licht en donker, daar wat richting in zien,
enzovoort, tot de diverse vormen van "oog" die er bestaan
.
Ogen zijn qua zenuwstelsel het meest veeleisende waarnemingsorgaan. Al was het maar
omdat er daar twee van zijn, en de signalen van de twee met groot voordeel
gecombineerd kunnen worden, tot een dieptebeeld. Waarvoor wel druk gerekend
moet worden - om één enkel beeld van de werkelijkheid te construeren. En
daarna het bewegingsapparaat te sturen om een pad af te leggen binnen die
geconstrueerde werkelijkheid. Een vorm van virtual reality zoals iedere
waarneming door wat dan ook een vorm van virtual reality is. Welke term hier
gebruikt wordt om aan te geven dat daar bijzonder veel rekenkracht voor
nodig is, want computers zijn pas een kort geleden krachtig genoeg geworden
om dit kunnen. Waarvoor in de hersenstam de eerste stappen worden gezet.
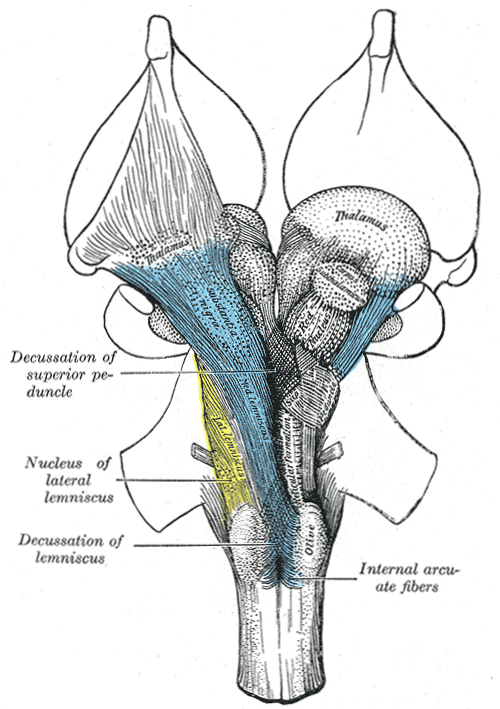
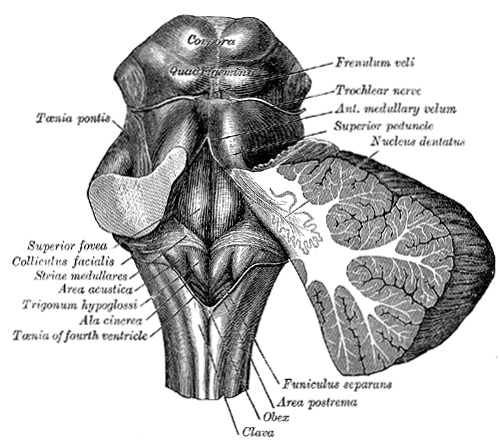
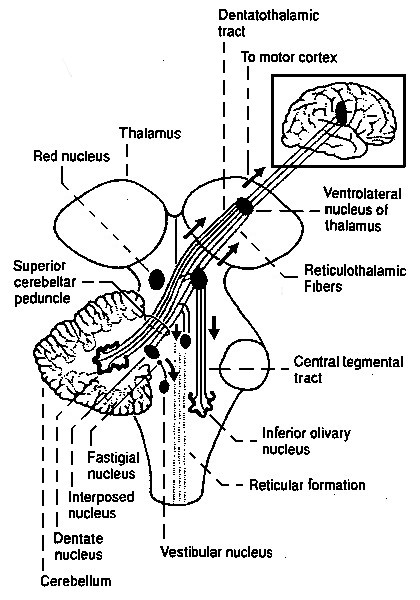
De volgende illustratie
geeft het eerste globale overzicht (voor/achteraanzicht) van de hersenstam - deze en de andere anatomische gravures
komen uit de atlas van Gray
- deze illustratie is Gray 690:
In deze gravures zijn de gestippelde structuren de kernen, concentraties van
neuronen, en de gestreepte zijn de bundels van neuronuitgangen of axonen die voor de in- en uitvoer van signalen
van elders zorgen bundel. Elders in de hersenen zijn de kernen meestal
duidelijker omlijnd, en redelijk bolvormig, in de hersenstam ligt dit wat minder
duidelijk, hetgeen aanleiding heeft gegeven tot een reeks van aanduidingen
als nucleus, corpus, locus, formation
enzovoort. Voor de bundels axonen is er een soortgelijke "verwarring" met termen als peduncle
(Latijn voor "stengel"), fasciculus ("bundel"),
lemniscus ("band") of gewoon "fiber".
De bundels vervoeren de informatie van de ene structuur naar
de andere. In de hersenstam lopen er bundels van en naar het ruggemerg, en
komende en gaande naar de hersenstamkernen waarvan de grootste is het hier getoonde complex genaamd de
olive of olivary body of olijfvormige lichaam, van
en naar de kleine hersenen of cerebellum waarvan hier in omtrek de
aan- en afvoer bundels zijn getekend net boven de olive, de
thalamus, het doorgeefstation naar de emotieorganen (waarvan hier ook zichtbaar
de caudate nucleus, de ui-achtige vormen aan de bovenkant - zie ook Emotie organen,
overzicht
 ), en direct van en naar naar de cortex
(niet weergegeven). ), en direct van en naar naar de cortex
(niet weergegeven).
Een ander belangrijk begrip dat hier weergeven wordt, is
decussation - dit is de algemene term voor "kruislings", wat wil zeggen dat
de bundels van de linker- naar de rechterhelft van het ruggemerg of de
hersenstam lopen en omgekeerd. Dit is ter coördinatie van de besturing van de linker- en
rechterhelft van het lichaam. Dit slaat hier op de Med. lemniscus of medial lemniscus
de bundel die
informatie uit de kernen van de hersenstam doorgeeft naar boven
, en de Sup. penducle,
een afkorting
voor superior peduncle, de bovenste van drie grote bundels naar het
cerebellum, waarover later meer.
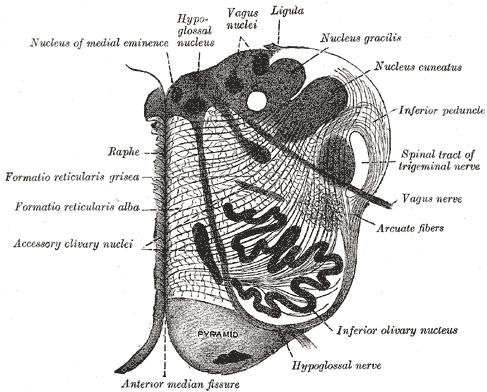
De eerste grote herkenbare
structuur in de hersenstam, kijkende van onderaf, is dus de olive. De olive
is ook de eerste met de gekronkelde lagenstructuur die terugkomt op diverse
andere plaatsen zoals het cerebellum en de cortex, zie deze horizontale
doorsnede (Gray 694):
Uit de vorm valt ook de soort functie af te leiden: het bestaan van een
zichtbare aparte laag van neuronen, of preciezer: neuron-kernen, valt af te
leiden dat er een specifieke functie wordt vervuld voor een conglomeraat van
andere neuronen, wier signalen binnenkomen in de laag middels bundels
axonen, uitgangen van andere neuronen (in anatomische secties zijn de
kernlagen donkerder en deze worden algemeen aangeduid met de term "grijze
massa" of "grey matter" en de gebieden met verbindingen zijn lichter
en aangeduid met "witte massa" of "white matter"). De uitkomst van de functie vertrekt
weer via de bundels van axonen van de laag zelf. Dit volgens het algemene
schema:
In eerste instantie ontstaat er een bolvorm omdat de neuronkernen meer
ruimte in beslag nemen dan de axonen, en groeit deze verhouding heel erg
scheef, dan ontstaat de noodzaak voor kronkels om alles in een beperkte
ruimte te persen, zoals ook bij de grote en kleine hersenen.
Bij dit simpele schema rijst de vraag: waarom de
functie niet vervuld ter plekke van de verzendende neuronen? Dat zou dus
vrijwel zeker ook gebeuren. De gebruikelijke praktische toepassing van het
schema is er dan ook eentje met twee ingangsbundels, en de voor de
hand liggende functie is dan: coördinatie. De volgende schematische
horizontale doorsnede illustreert dit met betrokkenheid van de olive (Gray
699, van hier
):
Dit schema centreert eigenlijk rond de loop van de onderste bundels van en
naar het cerebellum, de inferior peduncles (nummer 8). Maar
zichtbaar is dat een belangrijk deel ervan komt van de olive. Ook zichtbaar
is dat de olive signalen ontvangt van kernen 5, 6, en 7, de gracilis,
cuneatus en cinerea kernen, ontvangers van
proprioceptische en fijne-aanrakingsinformatie afkomstig van het ruggemerg.
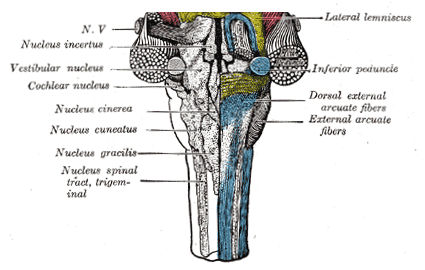
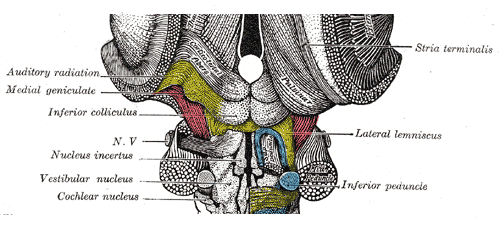
Ook in de buurt en zichtbaar in de volgende illustratie (deel van Gray 691,
een achteraanzicht met rechts de buitenste structuren verwijderd) zijn de kernen die de
signalen afkomstig van oor en evenwichtsorgaan ontvangen: cochlear nucleus
en vestubular nucleus:
De olive ligt dus het meest naar beneden dus het dichtst bij het
primaire bewegingsapparaat, en het ontvangt signalen over lichaamsstand en
waarnemingsignalen. Het ligt voor de hand om te veronderstellen dat hier de
eerste meer algemene coördinatie plaatsvindt tussen die signalen van het
waarnemingsapparaat (het N.V. is een afkorting van nervus vagus,
een van de twaalf hoofd-zenuwbundels lopend naar het lichaam dat het
bestuurt en die allemaal ongeveer hier binnenkomengaande, deze gaande naar keel, borst en omgeving).
Een aanname die versterkt wordt door het feit
dat de olive op zijn minst een deel
van het uitgaande signaal naar het cerebellum stuurt, waarvan allang bekend is (uit
gevallen van van beschadiging) dat het zorgt voor het leren en aansturen van
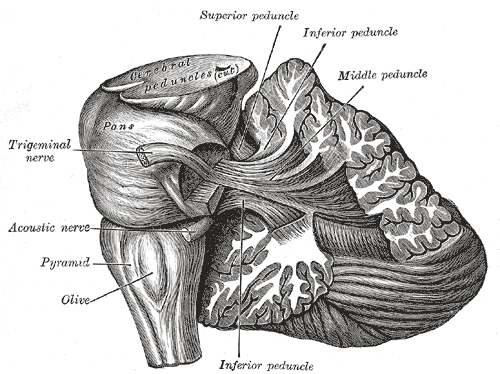
de fijne bewegingen. Hier een illustratie (Gray 705) op min of meer dezelfde
plaats, een kwartslag
gedraaid ten opzichte van de vorige:
Deze illustratie laat duidelijk zien dat de olive, ook van buiten, de eerst
zichtbare aparte structuur is (dit onderste deel van de hersenstam heet
medulla oblongata), gevolgd door de pons (of "brug"), die
leidt via de dikke bundels (peduncles) naar het grootste: het
cerebellum. Ook hier getekend de trigeminal of drielingzenuw, de hoofd-zenuwbundel
lopend naar het gezicht.
Is tot nu toe nog vermist één essentieel aspect van het zenuwstelsel
in zijn meest algemene ontwikkeling: het oog. Of beter in algemene termen:
het zicht, want in de loop van de evolutie zijn er vele soorten ogen
ontstaan. Uit hoofde van evolutionair voordeel meestal gelokaliseerd aan de
voorkant in wat werd een "hoofd". Vanwege de rijkdom aan informatie die het
verschaft een uiterst belangrijke ontwkkeling waaraan in primitievere soort
een flink deel van de hersencapaciteit was gewijd, dan genoemd het "tectum".
Dat is ook bij de mens, waar het relatief veel kleiner is, nog aan de
buitenkant zichtbaar (Gray 709, een kwart slag gedraaid ten opzicht van de
vorige, en verder naar boven lopend):
Het tectum is hier aangegeven als
corpora quadrigemina ("de vier lichamen"), vier van buiten zichtbare
bobbels op de hersenstam. Ze worden gewoonlijk verdeeld in onderste twee: inferior colliculi,
en bovenste twee:
superior colliculi. De onderste twee ontvangen de bundels komend van het gehoorsysteem, de
bovenste van het zicht - de bolvorm weer veroorzaakt door de neurologische
laagvorming. Hier komt dus de oog-informatie in het spel aangaande sturing
van beweging. Ook die informatie moet natuurlijk naar het cerebellum.
Het cerebellum is dus veruit de grootste structuur in deze regio, en de op een na
grootste van het zenuwstelsel na de cortex. Omdat er ook een interne gelijkenis
is, noemt men het in het Nederlands ook wel de kleine hersenen, hoewel hun
functie sterk verschilt. Die structuur is, het best zichtbaar in de bovenste
illustratie, die van een groot vel dat in elkaar gekronkeld en gevouwen is,
kennelijk om zo veel mogelijk vel in zo weinig mogelijk ruimte te proppen,
met bijbehorende boom-achtige structuur van verbindingen naar en van dat
vel. Het vel is grotendeels homogeen - op alle plaatsen van ongeveer
dezelfde structuur. De in- en uitgang naar buiten verloopt grotendeels via
een viertal in het midden liggende kernen, waaraan dus de peduncles
vastzitten.
De functie van het cerebellum is dus het leren van de
belangrijke fijnere soorten van beweging. De grootte ervan is vermoedelijk
het gevolg van de voortdurende strijd tussen de soorten om zo veel mogelijk
uit elkaar buurt te blijven of op te eten - of anderszins voordeel te
behalen bij het uitvoeren van beweging. Meer over het cerebellum hier
.
Het cerebellum maakt natuurlijk ook gebruik van de input van de
waarnemingsorganen. Direct uit het ruggemerg komen de signalen van de meest
primitieve: tast en pijn. In de hersenstam komen de signalen van oor en oog
binnen, via de zenuwen, het oorsignalen bestaan uit twee delen: gehoor en
evenwicht. Die signalen moeten gecombineerd worden met de lichaamspositie en
-bewegingen. Dit gebeurt in een reeks van kernen, en dus vermoedelijk
volgens de methode stap-voor-stap: telkens de combinatie van twee factoren,
waar de volgende dan weer mee gecombineerd wordt.
Van het hoor-systeem
zijn we in Gray 691 al tegengekomen de cochlear nucleus, de eerste kern voor het gehoor, en de
vestibular nucleus, die van het evenwichtorgaan. Verder naar boven in
Gray 691 is ook zichtbaar de inferior colliculus
, die ook
geassocieerd is met het gehoor, zie een tweede uitsnede hieronder:
En daarmee is het niet gedaan, want ook in deze illustratie zijn aangegeven
de medial geniculate en auditory radiation. De medial
geniculate of meestal medial geniculate body of medial geniculate
nucleus
genoemd, is een
kern verbonden aan de thalamus
, de structuur die de brug vormt tussen hersenstam
en emotie-organen en cortex - in de rechterhelft is de thalamus aangeduid met
pulvinar, de naam voor de achterkant ervan (de twee helften van de
illustratie vertonen verschillende niveaus van afpellen van de hersenstam).
De auditory radiation is de bundel die
uitwaaiert van de thalamus naar het gehoorsgedeelte van de cortex. Alle tezamen
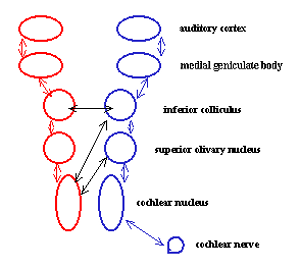
krijg je dit schema
:
In de hersenstam zelf zien we dus al drie opeenvolgende kernen: degene
waaraan de zenuw vastzit, degene die in de buurt van de olive, en de inferior
colliculus. De tweede combineert vermoedelijk het geluidssignaal met bekende
oorposities aan de hand van lichaamsposities. Hier kan dan ook het
richtingsbeeld ervan ontstaan. De inferior colliculus zou dan hogere vormen
van combinatie of filtering kunnen doen.
Daarna gaat het signaal naar de
medial geniculate
body van de thalamus, in welke laatste naar vermoeden de eerste integratie
van het totale lichaamsbeeld-plus-waarnemingsbeeld van werkelijkheid,
een virtual reality, wordt gevormd.
Het tweede
complexe waarnemingsystemen is dat van het zicht. De informatieverwerking die start met het menselijke oog is de uitkomst
van de informatieverwerking van de primitievere vormen van zicht. Die is
globaal vermoedelijk verlopen als volgt: licht = energie = groei = voedsel =
gunstig, dus: meer licht = eropaf. Maar snel meer of minder licht =
(vermoedelijke) beweger = kans op roofdier = snel wegwezen. Dit is dermate
belangrijk dat deze splitisng al in de buurt van het oog plaatsvindt. In het oog zelf wordt
dus niet gewoon
informatie opgevangen en doorgegeven, maar vindt ook meteen al verwerking
plaats: de directe optische informatie is een grote hoeveelheid lichtpunten,
pixels in computertermen, hetgeen het netvlies al omvormt in contouren,
vlakken enzovoort - niet direct in de lijn van deze globale beschrijving
maar zeer instructief voor het begrip van de werking van het zenuwstelsel in
het algemeen is de hier
gegeven
beschrijving van de werking van het oog.
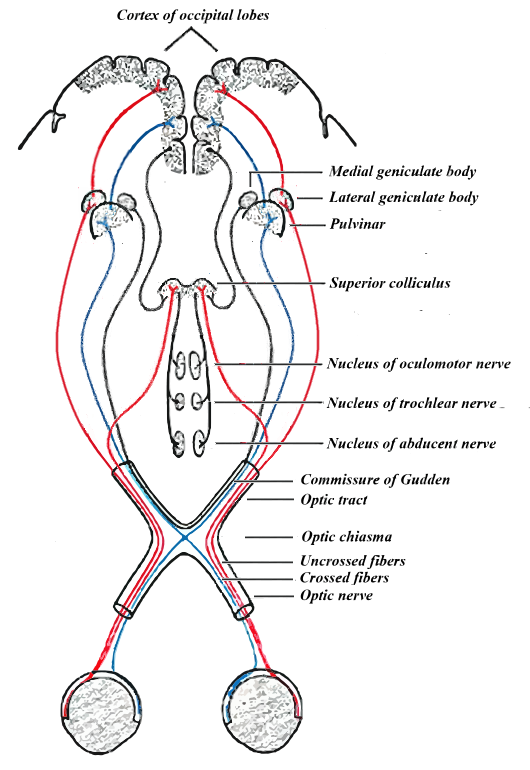
Vanaf het oog is de
ketenstructuur goed te volgen in de volgende illustratie van het
signaalverloop direct volgend op het oog - met de onderdelen in de hersenstam
in het midden (aangepast van Gray 722):
Eerst wat over de eigenaardigheden van de signaalverloop.
Helemaal onderin zijn weergegeven de oogbollen met achterin het netvlies en de
neuronuitgangen ervan, gesplitst in een linker- en rechter aandachtsveld apart
aangeduid met een rode en blauwe kleur. Deze scheiding is ook daadwerkelijk vastgesteld
bij mensen met trauma's aan specifieke onderdelen van het systeem, die soms
leiden tot zien in slechts één helft van het gezichtsveld.
Bovendien
is die scheiding in het ene oog ook nog eens omgekeerd ten opzichte van het
andere oog. Al die eigenaardigheden zijn in één klap te verklaren door te
constateren dat zaken dicht bij het oog voornamelijk in de rode gebieden
geprojecteerd worden, en zaken veraf dus meer in de blauwe. Dichtbij en veraf
hebben duidelijk geheel verschillende betekenis voor het organisme: dichtbij
vergt onmiddellijke attentie en potentieel snel handelen, dat wil zeggen:
afhandeling in de hersenstam - veraf moet gaan richting toekomstige scenario's
en vergelijking met vroegere scenario's, dat wil zeggen: richting emotie-organen
en cortex. De scheiding in dichtbij en veraf gebeurt op grond van de verschillen
in de beelden in linker- en rechteroog, dat wil zeggen: door middel van een vorm van "van elkaar aftrekken". Mede daarvoor is het
optic chiasma, dat alvast de banen mengt.
De volgende
fase is de verwerking in de hersenstam. Dat gebeurt in de superior
colliculus, zie Gray 691 boven. De superior colliculus ligt direct boven de
inferior colliculus, hetgeen tezamen met de gelijksoortige structuur,
een gelijksoortige integrerende functie doet vermoeden. Dit wordt ondersteund
door haar interne structuur, zoals geïllustreerd onder
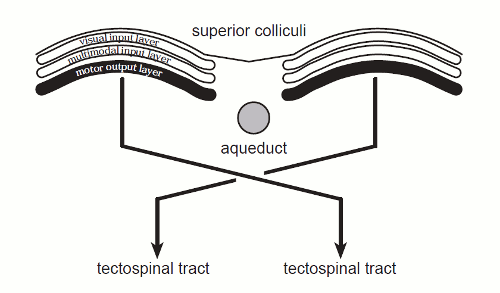
(van hier
- een schematische horizontale doorsnede - aqueduct is het vertikaal
lopende kanaal voor hersenvloeistof - tectospinal tract is hier de naam
voor de bundels van en naar het tectum bevattende de colliculi - ook zichtbaar is de zoveelste decussation, kruislingse verbondenheid):
Weergegeven is hoe de superior colliculus is opgebouwd uit lagen (ieder weer
bestaande uit meerdere onderlagen) - de visuele input wordt gecombineerd met
input van andere structuren (multimodal input layer), en derde laag heeft
verbindingen naar het ruggemerg (tectospinal tract),
hoogstvermoedelijk dus aansturing van bewegingen.
In het
keten-overzicht van Gray 722 zijn ook verbindingen vanaf de superior
colliculus naar de rest van de hersenstam zichtbaar - dit zijn de kernen die de
bewegingszenuwen van het oog aansturen. Wat kennelijk dusdanig belangrijk is dat
er aparte kernen zijn voor een paar aparte draai-spieren van het oog. De
trochlear nucleus bestuurt het naar beneden kijken en de draaiing om de
lijn van zicht (ter compensatie van het schuin houden van het hoofd). Een
speciale spier voor het naar beneden kijken heeft vermoedelijk dezelfde reden
als de links-rechtsspliting van het gezichtsveld: het bepaalt kijken naar
dichtbij of veraf. Ook het handhaven van de horizon is natuurlijk van eminent
belang. Wat de andere gespecialiseerde kern, de abducent nucleus, doet
kan men dan eigenlijk wel raden: die draait het oog naar links en rechts. De
overige functies van het oog worden geregeld door de oculomotor nucleus.
Het feit dat de besturingskernen en zenuwen vastzitten aan de
superior colliculus betekent dat de laatste op zijn minst mede bepaalt waar naar
gekeken wordt - dus bepaalt wat belangrijk is. Zoals iedereen kan weten in dat
bijvoorbeeld beweging in je ooghoek een bijna automatische reactie van "er naar
gaan kijken" veroorzaakt.
Een voorbeeld van de integratie van functies die in deze omgeving uitgevoerd moet worden, is het
combineren van de informatie uit het evenwichtsorgaan met de visuele waarneming.
Een schuin staand hoofd leidt in de waarneming niet tot een schuin staand beeld.
Dat is omdat het evenwichtsorgaan signaleert dat het hoofd schuin staat, en het
systeem van visuele waarneming die schuine stand verrekent in wat ze aan de rest
van het zenuwstelsel doorgeeft, zie de eerste bron hier
 . Deze berekening moet ergens gebeuren, en aangezien het een bijzonder
fundamenteel soort behoefte is tot bij de laagste diersoorten, gebeurt dit
waarschijnlijk ergens in de hersenstam.
. Deze berekening moet ergens gebeuren, en aangezien het een bijzonder
fundamenteel soort behoefte is tot bij de laagste diersoorten, gebeurt dit
waarschijnlijk ergens in de hersenstam.
Het tweede niveau in de zicht-keten zijn twee delen van de
thalamus: het lateral geniculate body, een aan de buitenkant liggende
kern ervan, en de pulvinar, de naam voor het achterste deel ervan.
De thalamus, zoals al gezien bij het gehoor, ligt zowel qua locatie als functioneel tussen hersenstam en
emotie-organen. Bij de thalamus komen alle waarnemingssignalen binnen, en is mogelijkerwijs dus
de eerste laag van totale integratie van de waargenomen wereld.
En als laatste laag in de
keten gaan zowel vanuit superior colliculus, lateral
geniculate body als pulvinar signalen richting de cortex, alwaar de zaak voor
het bewustzijn wordt verwerkt. Van de cortex gaan ook signalen richting superior
colliculus, voor de bewuste sturing van de ogen.
Meer over het proces
van integratie verderop.
In ieder geval al na de ontwikkeling van het oog maar vermoedelijk ruim
eerder ontstond
behoefte aan twee soorten beweging: de gewone, zeg voor beweging richting
plaatsen met voeding, en de snelle, bij het signaleren van gevaar. Waarbij de
laatste meer energie kost, dus zuinig mee moet worden omgesprongen en weer
snel tot rust gebracht indien niet meer nodig. Wat je het meest efficiënt
oplost met een enkel systeem met twee standen die kunnen omschakelen. En in
wat subtielere vorm: met geleidelijke tussenstanden.
Het standaardsysteem
van het zenuwstelsel is het activeren van het ene neuron door het andere of
juist het blokkeren ervan met twee neurotransmitters (is deze zin niet
duidelijk, raadpleeg dan dit
): respectievelijk glutamaat en GABA. Het in
snelle modus brengen gebeurt door een derde neurotransmitter: noradrenaline
(of in Engelstalig gebied: norepinephrine) en het tot rust brengen door
serotonine. Deze laatste stoffen komen van voor dat doel bestaande neuronen
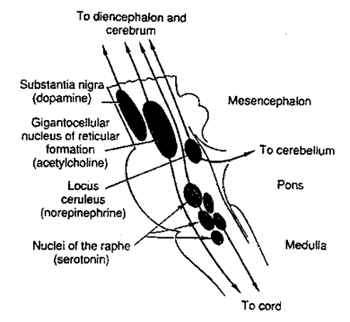
verzameld in specifieke locaties. Nordrenaline-afgevers in de locus
coeruleus (of "blauwe plek") en serotonine in meerdere raphe nuclei
(of "rand kernen")
(Wikipedia) - hier een globaal overzicht van hun locatie in zijaanzicht (van de site
van Ben Best
):
De raphe nuclei lijken tezamen hier veel groter, maar dat is niet zo want ze
zijn relatief dun, liggende aan de rand van de hersenstam, zie deze
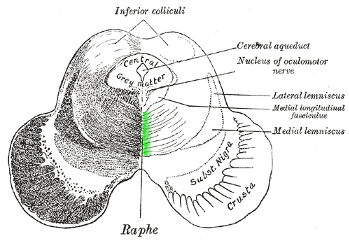
horizontale doorsnede (Gray 711):
"Raphe" slaat hierin op de scheidingswand tussen linker- en rechter
ruggemerghelft - de raphe nuclei liggen tegen die rand, zoals rechts
aangegeven met groen. De getekende lijntjes zijn de axonen die vanaf de kernen
naar de aan te sturen gebieden lopen.
De onderste raphe kernen
in het zijaanzicht sturen het
ruggemerg aan, die in het midden de hersenstam, en de bovenste de rest van
het brein.
Overigens: deze spreiding en randligging
lijken erop te wijzen dat het proces van het synthetiseren van serotonine
belastend is voor de neuronen in de omgeving.
De
onderste raphe-kernen spelen vermoedelijk ook een rol in de pijnafhandeling - zie bijvoorbeeld de
beschrijving van de functie van de nucleus raphe magnus
. Merk op: ook dit is al geen eenvoudige functie: pijn is zo belangrijk dat
het direct moet worden doorgegeven - maar in vluchtsituaties juist weer
niet, want ontsnappen gaat voor. Probeer een computerprogramma te schrijven
dat de betreffende sensorische gegevens ontvangt, en de juiste beslissing
neemt - dat gaat niet zomaar lukken met computerlogica en -programmatuur. De
natuur lost dit soort dingen anders op, is inmiddels bekend: middels neurale
netwerken
.
Uit hoofde
van compactheid en simpelheid is het dan het meest voor de hand liggend dat
dat in dezelfde buurt moet liggen. Daarvoor is er een kandidaat die normaliter
wordt beschreven als "hebbende een onduidelijke functie": de reticular
formation
(Wikipedia), of "gebied met netvormig uiterlijk"
- dat netvormig uiterlijk
zijnde een combinatie van lichte horizontale zowel als vertikale
streepvorming - zie nogmaals Gray 694 voor de doorsnede:
En nogmaals Gray 690 (onderste helft) voor het zijaanzicht:
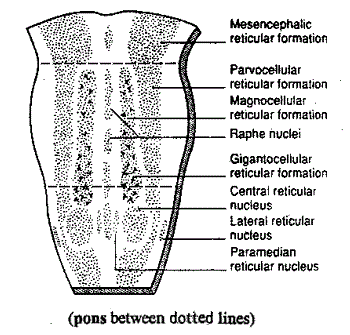
De reticular formation is dus een kolom die in het centrum van de hersenstam
loopt, gedurende de hele lengte. Met erin liggende de diverse kernen, en aan
de scheidingswand van links en rechts de raphe kernen. Zie dit schematische
voor/achteraanzicht in doorsnede (van de site van Ben Best, voor een grotere versie zie aldaar
):
De bovenkant (mesencephalon) sluit aan op het diencephalon (thalamus en omstreken) en de onderkant op het ruggemerg
(voor de encephalon-terminologie, zie hier
).
Voor
de activerende neurotransmitter: noradrenaline of norepinephrine, gelden
vermoedelijk dezelfde overwegingen als voor serotonine, zij het moeilijker zichtbaar omdat de
locus coeruleus niet zo handig verpreid is zoals de raphe kernen.
Dit
gaat dus over het schakelen tussen gewoon- en noodgedrag. Iets dat deels
aangeboren en deels aangeleerd wordt
. Daarmee is ook een oordeel
over de afloop en daarmee de wenselijkheid van bepaalde gedragingen
geïntroduceerd.
Combinaties van waarneming en gedrag die wenselijke resultaten opleveren
(meer voedsel) moeten een positieve waardering krijgen, en iets dat leidt
tot achternagezeten worden door een roofdier een negatieve. Dat oordeel is
neurologisch niets anders dan het bestendigen van bepaalde neuronale
verbindingen en het loslaten van andere. Ook dat gaat dus met
neurotransmitters.
Dit zijn qua besturingstechniek hogere functies en
geheel toepasselijk liggen de bijhorende
structuren dan ook weer hoger in de hersenstam: de bestendigende en doen-herhalende
neurotransmitter is alom bekend als dopamine, en de vermijdende minder
bekende (niet-gedrag is lastiger waar te nemen) dus acetylchlorine, want dat
is de vierde neurotransmitter die in de hersenstam wordt opgewekt.
De dopamine-neuronen
zijn gelokaliseerd in een gebied genaamd als VTA, oftewel ventral tegmental area. De term "tegmentum" wordt meer gebruikt en hoort bij tectum volgens
onderstaande illustratie (tezamen vormen ze dus de bovenkant van de hersenstam of ook wel midbrain
of mesencephalon):
Het ventrale gedeelte is het "buikliggende" (naast "dorsaal" of
"vinliggend"). In onderste versie van het eerder gegeven zij-aanzicht is
het tegmentum groen, en het ventrale gedeelte met gele arcering erin:
De dopamineproductie ligt daarin gecentreerd in het deelgebied ervan
genaamd substantia nigra (- pars compacta) - merk op: ook
dit is een vrij plat gebied, dunner dan hier getekend, liggende tussen
vertikaal lopende bundels verbindingen.
De acetylcholine-gebieden
liggen lager en meer aan de buitenkant, voornamelijk in de pedunculopontine nucleus
of gigantocellalur nucleus.
De combinatie van dopamine en acetylchlorine-gebieden en het omliggende
deel van de reticular formation en mogelijk nog wat andere structuren vormen
ook wat wordt aangeduid als het "reticular activating
system"
- de structuren die het niveau van alertheid en de overgang tussen waken en
slapen bepalen. Die laatste overgang gaat, zoals bekend, gepaard met het
volledig loskoppelen van het bewegingsapparaat, waarvoor dus ook een
gelokaliseerde structuur moet bestaan die deze "schakelaar" overhaalt. En het
bestaan van het verschijnsel van "flauwvallen" laat zien dat beide processen ook gelijktijdig
kunnen worden geactiveerd. Voor de uitvoering hiervan worden ook weer de modulerende neurotransmitters
gebruikt, zoals in extrema getoond wordt
door ze uit te schakelen, wat is gedaan met dierexperimenten. Van de site van Ben Best
:
Bekend is dat de basale schakelaar die de overgang naar slaap bepaalt ligt
in de hypothalamus (zie verder), maar deze experimenten laten zien dat de
uitvoering van de opdracht ligt in de hersenstam. Met de modulerende
neurotransmitters als daadwerkelijke uitvoerders. Meer over slaap hier
.
Dit voorlopig wat betreft de aansturende biochemie van het basale
neurologische systeem, waarin van beneden naar boven is gewerkt. Ondertussen
zijn een aantal onderdelen overgeslagen vanaf het cerebellum.
In de
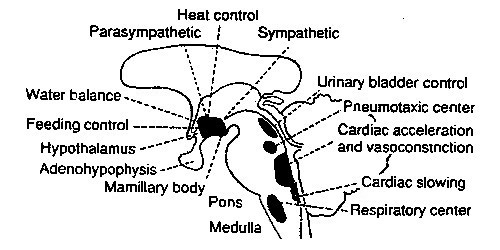
pons en verder naar boven ligt nog een reeks kernen,waarvan een deel het
autonome zenuwstelsel aantrut, zoals al weergegeven in het schema van het
ruggemerg. Dit gaat over het autonome zenuwstelsel, de "huishoudelijke" functies gekoppeld aan de organen:
hartslag, bloeddruk, slaap, ademhaling, slikken, blaasbesturing, enzovoort. Ook dit
wordt dus grotendeels afgehandeld volgens de aanpak: "per functie een eigen kern",
hoewel de "kernen" een vrij losse structuur hebben - zie
onderstaande overzicht (van de site van Ben Best
, met links-rechts oriëntatie aangepast aan de standaard van deze site):, met links-rechts oriëntatie aangepast aan de standaard van deze site):
Aangeduid in deze tekening is ook de opsplitsing van het autonome zenuwstelsel
in een (ortho-)sympathisch en para-sympathisch deel, (ortho-)sympatisch staande
voor de activerende functies, de reacties op het buitengebeuren, en para- voor
de functies in ruste, zoals de spijsvertering.
Maar hieronder zijn dus
al functies die duidelijk wel beïnvloed kunnen worden door het
bewustzijn zoals de blaascontrole.
Dit wijst er dus op dat er naar
boven toe steeds meer integratie en coördinatie plaatvindt. Een kern hij
niet benoemd is degene die het voedsel-vermalen aantuurt, in het Engels:
mastication. Het ontwikkelen van een aparte structuur voor dergelijke
functie, wat natuurlijk energie kost, is kennelijk de moeite waard. Met als
eerste zichtbare factor: de aandacht: de rest van het brein kan een opdracht
sturen naar deze kern: "kauwen", en daarna zijn aandacht op andere dingen
richten. Dergelijke kernen zijn dus te beschouwnen als vormen van
"programmatuur": er wordt een reeks meer elementaire opdrachten doorlopen,
zoals "kaak omlaag - voedsel manipuleren met tong richting kiezen - kaak
naar beneden" enzovoort.
Het is voor de hand liggend om te
veronderstellen dat in de gang van onder naar boven in de hersenstam, en
mogelijk ook al in het ruggmerg, voortdurende sprake is van het samenvoegen
en coördineren van steeds gecompliceerdere combinaties van meer elementaire
bewegigngs- en waarnemingsfuncties. Met structuren die het kenmerk hebben
van programmatuur: het aflopen van vaste patronen van beweging en
bijbehorende zaken: geef een impuls aan het begin van het netwerk, en het
netwerk loopt door een aantal stappen heen die andere onderdelen aansturen.
Nog een paar structuren van de
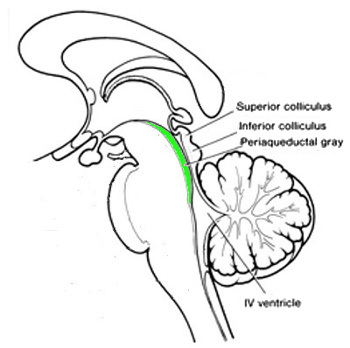
hersenstam verdienen aparte vermelding. Als eerste het peri-aqueductal grey of
PAG
, weergegeven hier:
Het is Engels-Latijn voor "rond de waterweg liggende grijs", de waterweg
zijnde de verbinding voor hersenvloeistof van boven naar de IV ventricle
waarin "ventricle" is "ruimte met hersenvloeistof" - de hersenvloeistof is voor
koeling, schokopvang en dergelijke. Het "grijs", zoals gezien betekent dat het
voornamelijk bestaat uit neuronkernen en onderlinge
kort-afstandsverbindingen middels hun dendrieten.
Het is
bekend in de neurologie dat het PAG een rol speelt bij het doorsturen van
pijnsignalen naar het bewustzijn. Onafhankelijk daarvan is uit het gewone
leven ook bekend dat onder speciale omstandigheden de pijnsignalen volledig
geblokkeerd kunnen worden. En uit andere culturen is bekend dat het
bewustzijn ertoe getraind kan worden om pijnsignalen te negeren, oftewel te
blokkeren, en de PAG krijgt inderdaad ook signalen vanuit de cortex. Dat
alles wijst erop wijst erop dat er een specifieke plaats is waar beslist
wordt de pijnsignalen worden doorgegeven, en dat het PAG die plek is.
Dan nog twee structuren die normaliter bij het limbische systeem
of de emotie-organen
worden ingedeeld:
hypothalamus en hypofyse (Engels: pituitary gland of
pituitary), en bijbehorend de epifyse. Eén argument is al besloten in
tekeningen als bovenstaande, die de evolutionaire structuur van de hersenen
volgen, en die alles horende bij de hersenstam automatisch bij elkaar
houden:
Met links de hypofyse en rechts de epifyse. Die zijn tezamen bekend als
de klieren ("glands"). Wat ze doen, is neurotransmitters in het bloed
pompen, die dan hormonen worden genoemd, om de boodschappen van de hersenen
ook algemener in het lichaam te verspreiden. Dit is natuurlijk al essentieel
voor de functies uitgevoerd door de hersenstam.
Nog een goede reden
voor herindeling: gebruik je het criterium dat alles wat in zichtbare
links-rechts variant komt hoort bij emotie-organen en hoger, en de
enkelstuks bij de hersenstam, dan is dit ook een reden want van hypofyse,
epifyse en hypothalamus is er maar één.
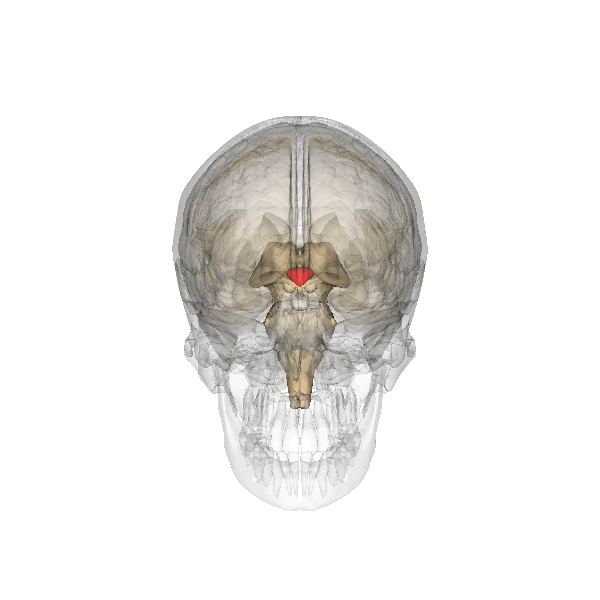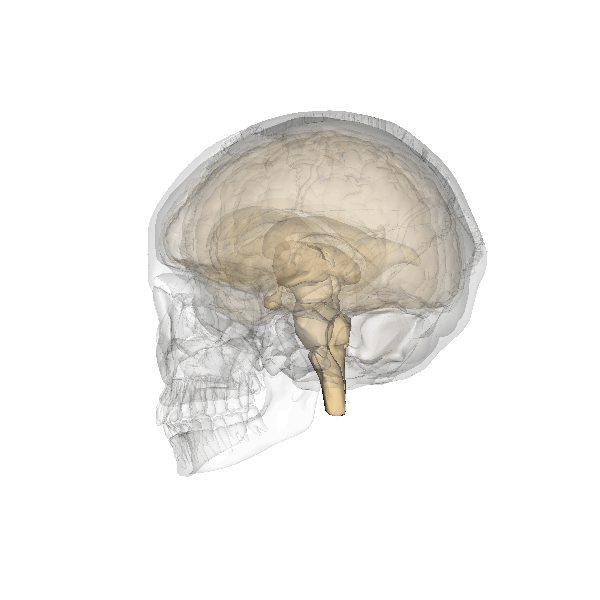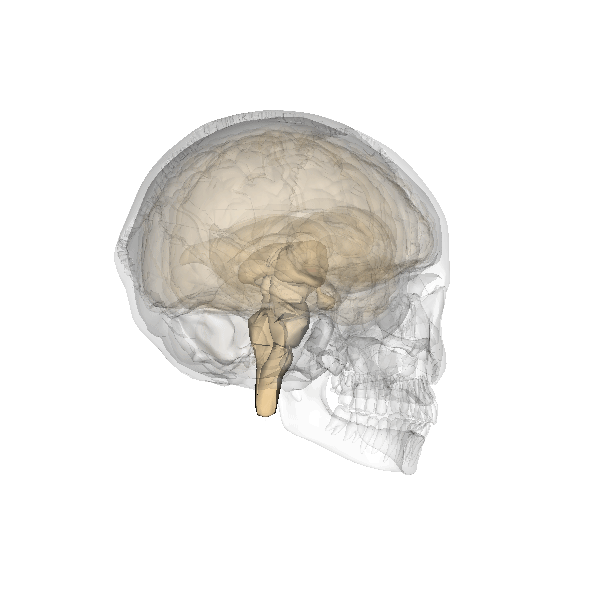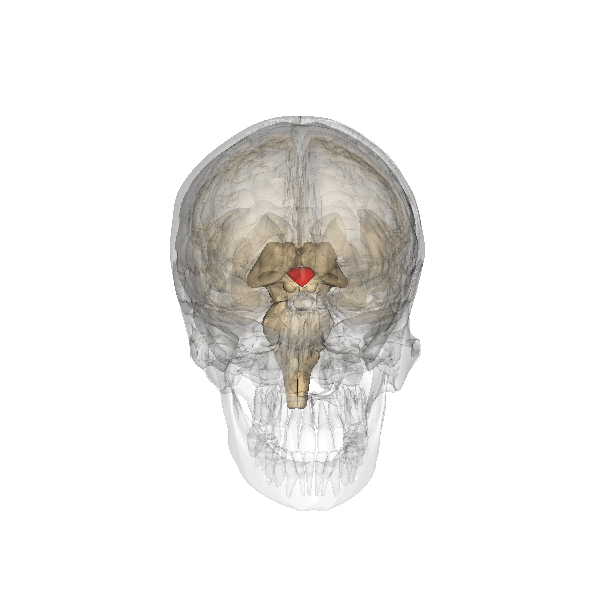
De hypothalamus en haar juiste locatie is lastig uit de beelden in twee
dimensies, vandaar deze:
De hypothalamus is het roodgekleurde element, en de hypofyse is het grijze
'bolletje" dat eraan vastzit.
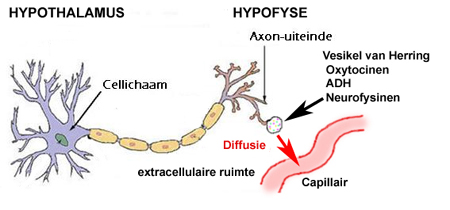
De klieren zijn de enige organen die stoffen direct kunnen transporteren tussen
lichaam en hersenen (die zijn biochemisch strikt gescheiden, in verband met
infectiegevaren), de rol van de hypothalamus daarin is die van aanstuurder, zie de volgende illustratie:
De functies van de hypothalamus zijn af te leiden uit de lijst van verbindingen
ernaar
(Wikipedia, opgeslagen 02-04-2012):
Of uit de Nederlandse versie:
Uit welke lijst met functionaliteiten je ook onmiddellijk de conclusie kan
trekken dat de hypothalamus en hypofyse functioneel eerder bij de hersenstam dan
bij de emotionele organen horen.
Ook is hieruit duidelijk dat de hypothalamus en hypofyse een faciliterende
functie hebben - de reden voor het afscheiden van hormonen wordt bepaald elders.
De hypothalamus speelt dus ook een essentiële rol in nog een basale
functie van het brein: de slaap. In de hypothalamus zit de schakelaar die de
overgang naar slaap in gang zet - ook is ze de
bron is van de bijbehorende basale neurotransmitter histamine.
Inbouw
Dit wat
betreft het functioneren van de hersenstam als min of meer zelfstandig orgaan.
In de mens (en in ieder geval ook de zoogdieren) wordt de functionaliteiten van
de hersenstam deels aangevuld en deels overgenomen door ten eerste het
emotionele znuwstelsel en ten tweede de cortex. De hersenstam werkt dus met deze
twee samen, en daar gaat het nu verder over. Als eerste de
neurotransmitter-koppelingen en daarna worden de subsytemen
horende bij horen en zien als voorbeeld genomen. En tenslotte het hele
bewegingssysteem.
De eerste koppeling wordt weergegeven in overzichten waarin de
hele hersenstam is uitgebeeld tezamen met hypothalamus en/of hypofyse, zoals
boven een paar en hieronder met groen is benadrukt:
Het groene gebied staat voor een bundel van verbindingen van hersenstam naar
hypothalamus, noodzakelijk voor het gebruik van de hormonen door de hersenstam.
De tweede en derde reeks komen voor in een andere reeks schematisch
afbeeldingen, worden daar ook zelden tot nooit benoemd, en zijn elders ook
moeilijk vindbaar:
De linker bundel (ze nemen niet het hele gebied in beslag, maar de bundels
liggen in dit gebied) is het mammilotegmental tract
, dat wil zeggen: verbindingen tussen hersenstam en mammilary bodies,
behorende tot de emotie-organen, en vermoedelijk betrokken bij gedragsevaluatie
en geheugenvorming.
De rechter staat voor een bundel verbindingen
lopende van de habenula
of habenular nucleus naar de
interpeduncular nucleus
en het tegmentum, de locatie van de neurotransmitter-organen (SN, LC enzovoort).
Ook de habenula
speelt een rol in het proces van ervaringsverwerking en geheugenvorming.
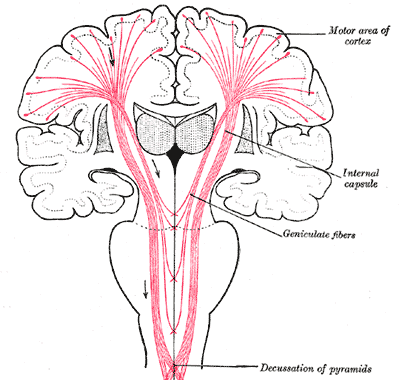
De derde reeks koppelingen van de hersenstam is van de lange-afstand soort,
en wordt meestal weergegeven tezamen met de directe verbindingen van ruggemerg naar cortex -
in "het grote schema" van het brein,
zoals in onderstaande illustratie uit de reeks van Gray (nr. 764):
De internal capsule is de plaats waar de bundels de dichtbevolkte
omgeving van de emotie-organen doorsnijden - de twee bollen moeten de twee
thalamussen voorstellen, de naburige grijze driehoeken onderdelen van de basale
ganglia.
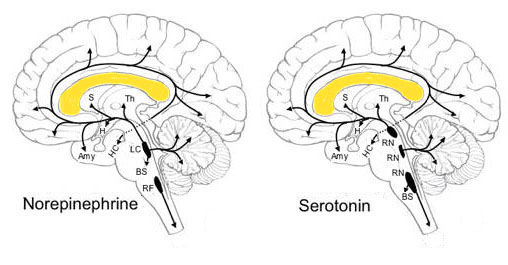
De vierde reeks verbindingen is zichtbaar in de
reeks illustraties van de neurotransmitterbronnen naar de rest van het brein. De
vier modulerende neurotransmitters worden opgewekt in de hersenstam maar
gebruikt in het hele brein inclusief de cortex. Dat betekent dat er paden van
bundels van uitgangen van de donerende neuronen naar het hele brein lopen -
onderstaande illustratie laat er twee zien
(het origineel is gewijzigd naar de hier gebruikelijke oriëntatie voor een
zijaanzicht: met de neus links, en wat cosmetische correcties aangebracht), maar
de overige twee hebben hetzelfde patroon:
| |
RN: raphe nuclei; BS: brain stem (hersenstam); Th: thalamus; S: septal nuclei; HC:
hippocampus; Amy: amygdala; H: hypothalamus; LC: locus
ceruleus; RF: reticular formation.
De hypothalamus ligt direct onder de H - het uitsteeksel onder de H is
de hypofyse. De hippocampus is aangegeven als bestemming maar
niet zelf zichtbaar. De aanduiding RF als bron van noradrenaline
voor het ruggemerg is correcter LTF of lateral (zijwaartse)
tegmental field. |
|
Dit is slechts een schematische weergave - in werkelijkheid ligt de wijdste
boog bovenop de hersenbalk (aangegeven met geel) in een laag van verbindingen,
het cingulum
, tussen de hersenbalk en de eerste laag van de cortex, de cingulate cortex -
dit geldt voor alle vier neurotransmitters. Dat de linker en rechter
afbeelding vrijwel identiek zijn, is dus geen luiheid - het is de
werkelijkheid. Hier een fMRI -opname van de verbindingen vanaf het cingulum
de cortex in:
Het dunne cingulum lijkt hier dikker omdat dit een driedimensionale opname
is. Merk op dat dit de werkelijkheid is, en alle getekende schema's, vooral
van de verbindingen, meer functioneel dan daadwerkelijk zijn - hetgeen hier
net boven is nog één van de meer accurate.
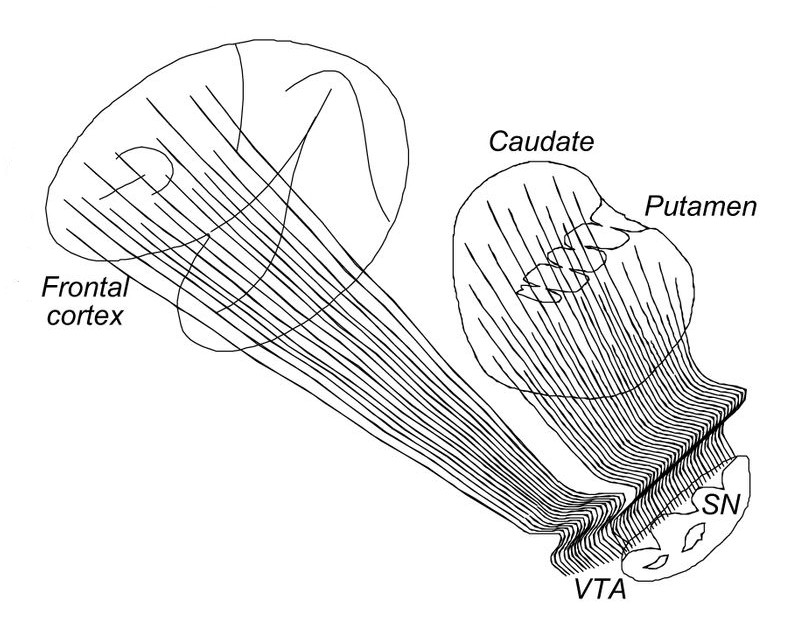
Het vierde voorbeeld van
de koppeling van systemen van hersenstam en de rest van het brein is dat van het
bewuste aansturing van het bewegingssyteem, en met name de rol van dopamine -
hier is het bijpassende overzicht
(let op: om beter aan te sluiten bij overige illustratie zijn rood en blauw
zijn omgewisseld):
Onderaan zichtbaar is de hersenstam met
daarin de substantia nigra (SN), in het midden de relevante onderdelen van de
basale ganglia
(de eerste laag
van de emotie-organen
, zie Emotie-organen, overzicht
), en dan de cortex die eigenlijk om het geheel heen ligt. Ook schematisch
getekend zijn de neuronale verbindingen: in blauw de activerende en in rood de
blokkerende.
De dikte van de pijltjes geeft hun relatieve sterkte aan, en in linker
hersenhelft is het
geheel min of meer in evenwicht. Rechts is het evenwicht verstoord, overeenkomende
met het optreden van de ziekte van Parkinson
.
De rol van dopamine hierin is dat het het evenwicht tussen de
activerende en de blokkerende processen regelt,
en de situatie in de rechter
hersenhelft is het effect van een tekort aan dopamine. De veelheid van
verschijnselen rond Parkinson bevestigt meteen dat dopamine geen
specifieke taak heeft, maar de algemene van moduleren.
In het rechtsstaande diagram van de circuits van verbindingen is één van de processen
betrokken bij Parkinson schematisch uitgewerkt (let op: dit is gewijzigd ten opzichte van het origineel
- om beter aan te sluiten bij de voorgaande illustratie zijn rood en blauw
zijn omgewisseld, en het diagram is geherordend naar neurologische locatie,
van beneden naar boven):
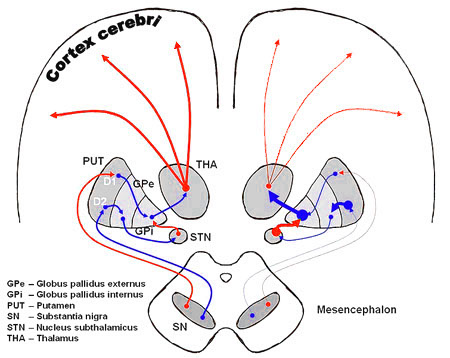
Hierin is striatum is de naam
voor de combinatie van putamen en caudate nucleus, twee van de
basale ganglia die liggen naast de globus pallidus (GPe en GPi), zie Emotie organen, overzicht
.
Merk als eerste op dat dit allemaal lus-circuits zijn, dus met
terugkoppeling
ingebouwd.
Merk als tweede op dat twee achtereenvolgende rode pijlen
gelijk zijn aan tezamen een enkele blauwe. Bijvoorbeeld in de combinatie
striatum - GPe - STN blokkeert het striatum de GPe, maar omdat de GPe de STN
blokkeert, zal een extra blokkade (uitgaande van een evenwichtssituatie) door het striatum neerkomen op minder blokkade
voor de STN.
Merk als derde op dat uit het striatum alleen blokkerende
signalen komen (overigens gaan de uitgangen van het striatum uitsluitend naar
andere onderdelen van de basale ganglia). Een mogelijke reden daarvan is dus
dat je uit twee opeenvolgende blokkerende signalen wel een activerend effect kan
krijgen, maar niet uit twee activerende een blokkerend (dit weerspiegelende
gelijksoortige regels uit de logica en de algebra: uit twee minnen kan je
wel een plus maken, maar van twee plussen krijg je weer een plus).
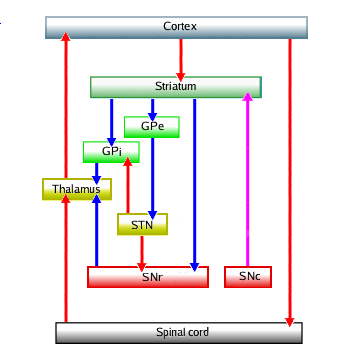
Uit het diagram zijn twee ondercircuits af te leiden: één dat het activatieniveau van de cortex verhoogt verhoogt, en één dat dat vermindert. En
het het gaat hier om de activatie van beweging. Het activerende, directe,
circuit is
:
Oftewel: cortex → striatum → thalamus → cortex gaat via GPi met twee maal rood
is netto één keer blauw, en via SNr met twee keer rood is netto één keer blauw,
oftewel: dit is beide meekoppelend.
Het deactiverende, indirecte, circuit heeft een extra stap via de STN,
In de stap via STN zit een extra rood plus een extra blauw, is netto één keer
rood, is een omkering. Oftewel: dit circuit is tegenkoppelend.
De normale toestand is dat beide circuits zich in een redelijk evenwicht houden, in het
"midden", met een gemiddeld activatieniveau horende bij normale menselijke activiteiten en
hun bijbehorende gematigde bewegingen.
Dit is het basale proces bestaande uit mee- en
tegenkoppelingen - het soort proces dat in Terugkoppeling
beschreven is als liggend onder het proces van het handhaven van evenwicht - een
stabiele toestand. De besturing van dit proces gaat door het verschuiven van dit
evenwicht van activeren en inhiberen. En daarvoor staat (mede) de tot nu toe niet besproken
paarse pijl lopende vanaf het SNc, het substantia nigra compacta, dat dopamine verstuurt
richting striatum. Dopamine stimuleert het directe circuit, en als van het
waarnemingssysteem of elders signalen komen die vragen om meer activiteit,
verstuurt het SNc meer
dopamine, en komt er meer activatie in de beweging. Met als een uiterlijk kenmerk dat de
persoon opgewonden raakt - en bijvoorbeeld gaat gesticuleren.
Bij ernstig gebrek aan dopamine raakt dit systeem, het
evenwicht, ontregelt, en een van de bekende gevolgen van verstoord
evenwicht is optreden van oscillaties (in een veer of iets dergelijks
mechanisch) of als het om mensen gaat: trillingen
, bekend als verschijnsel van de ziekte van Parkinson.
Als het omgekeerde van activatie nodig is, wordt het indirecte circuit
gestimuleerd, door een van de andere modulerende neurotransmitters: acetylcholine,
komende van wat lager gelegen kernen van de reticular formation, met name de
PPN. Tussen PPN en SNc liggen koppelingen die ervoor zorgen dat niet beide
stoffen tegelijk werkzaam zijn.
Dit is een overzicht waarin de details
ontbreken. Die details zijn onontbeerlijk voor het gaan richting toepassing,
waarvan één enkel voorbeeld. Onderstaand is een illustratie uit een
gereputeerde wetenschappelijke publicatie
maar (nog) niet elders gevonden:
Ze toont een fysiologische scheiding in het bereik tussen de twee
gebieden die dopaminebronnen zijn. Bekijk in dat licht de aanhef van de
volgende publicatie (de Volkskrant, 06-02-2017):
Oftewel: daar waar men een fysiologische scheiding ziet, vindt men ook
een functionele. Met, indien deze relatie bevestigd, grote consequenties
voor zowel inzicht in de werking van het geheel als de praktijk van
behandeling van disfunctioneren.
Conclusies De hersenstam vervult dus een paar soorten
fucnties naast zeer vele specifieke: de volkomen zelfstandige waaronder die
van de huishouding, een aantal waarin het het overige brein aanvult en een
aantal waarin het het overige brein, in geval van nood, vervangt. Het eerst
heeft het bewustzijn weinig tot geen invloed op en weinig mee te maken. Het
tweede leidt tot soms tot vaker onverklaarbaar gedrag, omschreven als
"menselijke driften" ern dergelijke. En ook een deel van het karakter. Het
derde zorgt in veel gevallen voor redding door snelle reacties daar waar het
bewustzijn de zaak niet zou kunnen bijhouden.
Dat laatste geval
wordt meestal omschreven als de situaties van "vechten of vluchten" - of nog bekender in het Engels als "fight or flight"
. Maar er zijn er nog twee: ten
eerste "bevriezen": niet-bewegen. In de praktijk onveranderlijk van de
variant "stokstijf", dus duidelijk een zelfstandige bewegings- en
neurale toestand. En dan is er nog
"aangetrokken worden". En het is meteen ook duidelijk waar dit nuttig voor is:
het verzamelen van voedsel en in extrema voor roofdieren het zien van een prooi,
en het verzamelen in groepen met soortgenoten, en het vinden van
voortplantingspartners.
Immobiliteit lijkt "niets" te zijn want je doet niets, maar is wel degelijk een
keuze-mogelijkheid van groot belang: voor talloze roofdieren is het signaal van
"een prooi" het bewegen ervan, en door bevriezen kan je je als poptentiële prooi aan detectie
onttrekken. En de situatie van niets-doen of bevriezen biedt de gelegenheid de
situatie te overwegen om tot een meer kansrijke reactie te komen.
Met een hersenstam die de meest essentiële beslissingen voor het
overleven neemt ook bij het bestaan van hogere hersendelen, is het de vraag
waarom die hogere hersendelen zijn ontstaan. De hoogstvermoedelijke reden is haar "digitale" karakter - het is "aan" of "uit" en weinig of geen
tussenfasen. Dat is een beperking en in veel omstandigheden een
tekortkoming.
De tweede, aanverwante, factor is die van de beperkte
leermogelijkheden: de overgangen van aan naar uit zijn te ingrijpend om door
ervaringen te veranderen, en vragen dan te veel ervaringen.
Daarmee komt een derde beperking om de hoek kijken: de beperkte capaciteit tot
het opslaan van ervaringen.
Met dat laatste is meteen een fundamentele
tekortkoming opgenoemd die dus eerst moest worden opgelost. Daarvoor dienen
vermoedelijk de eerste structuren van wat in de neurologie het
"limbische systeem" is gaan heten, en alhier wordt aangeduid met
"emotie-organen".
De emotie-organen kunnen subtielere beslissingen
nemen dan alleen "vechten enzovoort", aan de hand van overwegingen die de mens
dus "emoties" noemt. Wat een groot voordeel is maar ook een nadeel heeft:
die beslissingen duren langer om te nemen.
Een spel dat zich dus nog
een keer herhaalt met de rationele hersenen oftewel de cortex.
Deze beschrijving
kan dus gezien worden als een meer systematische uitwerking van het adagium van William
James
: "Je rent niet weg omdat je bang bent, maar je bent bang omdat je wegrent".
Waar de opmerking van James beperkt is tot de emotie van en de impuls tot
vluchtgedrag, wordt dat hier dus uitgebreid tot alle emoties die onderliggende
impulsen kennen (dat geldt niet voor alle emoties). Geformuleerd voor angst en
uitgebreid tot in de cortex: in de hersenstam is op grond van de waarnemingen besloten weg te
rennen, de emotionele hersenen interpreteren dat in het licht van voorgaand
gedrag en plakken daar het label aan dat bij dat eerdere gedrag is gaan horen, en dat label wordt gelezen door het
bewustzijn in de cortex: "Ik ben bang".
Waar het in deze beschrijving van de hersenstam mede om gaat, is om te laten
zien dat de hersenstam niet een volledig zelfstandig functionerende eenheid is
die voor de rest, met name het bewustzijn, de dagelijkse lichamelijke routine
afhandelt, maar een integraal deel uitmaakt van de stemmingen en sommige
beslissingen van het geheel van het zenuwstelsel, waartoe het bewustzijn ook
behoort.
Deze algemene invloed kan weer het duidelijkst geïllustreerd worden door de extreme
gevallen. Mensen doen soms dingen die anderen en ook henzelf totaal verbijsteren
- met als bekendste voorbeeld "zinloos geweld", maar er zijn meer soorten
. Waar het hier omgaat is dat ze zelf duidelijk geen controle op dit gedrag
hebben, dat wil zeggen: hun bewustzijn. Oftewel: de rationele hersenen. Ook de
emotionele organen zijn geen erg geschikte kandidaat, omdat de het bewustzijn
zich toch bijna altijd donders goed bewust is welke emoties er meespelen in
zaken. En bovendien worden vele van deze "impulsieve" daden gedaan op een
niet-emotionele manier.
De term "impulsief" zegt hier eigenlijk al alles: dit zijn
daden geïnitieerd door het reflexensysteem - de hersenstam. Dat dit gedrag
mensen verbijsterd, betekent dat ze niet weten waar het vandaan komt - omdat
het een laag te diep ligt om de oorsprong te kunnen waarnemen.
Dit zijn de extreme gevallen. Maar volgens het principe van de glijdende schaal
is het dus niet onaannemelijk dat ook bij vele andere beslissingsprocessen die
volledig door het rationele verstand genomen lijken worden, de hersenstam een rol
speelt. Die externe rol is op het psychologische vlak bekend voor de emotionele organen: het is
zelfs zo dat de meeste mensen in de meeste omstandigheden hun emotionele
ingevingen laten prevaleren boven hun rationele
. En als dat geldt voor een
(ruime) meerderheid van beslissingsprocessen, is het beslist niet onaannemelijk
dat, gezien de vrij intensieve koppeling tussen emotionele organen en
hersenstam, bij een deel van die beslissingsprocessen weer nog verderop van de
hersenstam stamt, waarvan psychologische voorbeelden hier
.
Die zijn het gevolg van neurologische koppelingen. Veel op het rationele vlak
onverklaarbaar gedrag, is wel verklaarbaar door ook de rol van de hersenstam in
het hele proces te betrekken
. Het is de vermoedelijke neurologische basis van verschijnselen als cognitieve
ziekte
en cognitieve psychopathie
.
Vervolg
Dit wat betreft dit overzicht van het functioneren van de hersenstam.
Ter afsluiting nog wat over de overige structuren aan de top ervan en het
overgangsgebied naar de emotionele organen. Waarvoor we weer teruggaan naar het
eerste overzicht (Gray 690):
Dit overzicht bevat ook de eerste structuren die niet tot de hersenstam horen.
De red nucleus
ligt net over de grens tussen mesencenpahlon (middenbrein) en dienencephalon
(dubbelbrein), waar het brein zichtbaar in twee helften is gesplitst, en heeft
vermoedelijk ook nog een directe bewegingscoördinatie functie, zie ook
onderstaand zijaanzicht:
En wat meer detail in dit achteraanzicht (van hier
):
De rode kern wordt in de literatuur bij dieren in verband gebracht met
bewegingsgang of tred ("gait"), en bij de mens met
bewegingscoördinatie in de babytijd, zoals kruipen.
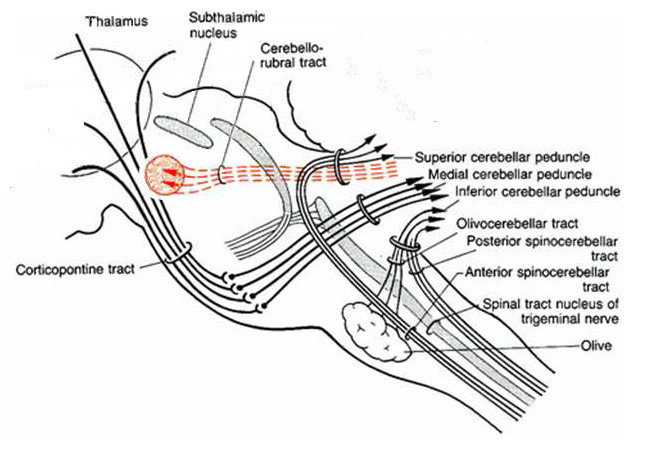
Ook ingetekend
in de eerste is de subthalamic nucleus (STN) - die is in interactie met de
globus pallidus en omliggende onderdelen, boven ontmoet in het
bewegingscircuit naar aanleiding van de rol van dopamine. Aldaar is de STN
de bron van de rode pijlen, oftewel exciterende neuronverbindingen,
terwijl de globus pallidus en omliggende onderdelen alleen verbonden zijn
met blauwe, blokkerende, verbindingen.
Nu zou je kunnen denken: dan
doet de globus pallidus enzovoort niets, maar dat is natuurlijk onzin. Ze
hebben kennelijk een statische functie, en de activerende signalen komende
van de STN zorgen ervoor dat ze die functie vervullen. Meer daarover bij de
emotie-organen.
Dan volgt de thalamus
zelf, die gezien wordt als het het algemene doorgangsstation van de "resultaten"
van de hersenstam. De thalamus dient mogelijk ook voor de allereerste algehele
integratie van het interne en extrne waarnemingsbeeld van het systeem. Er loopt
een belangrijk kanaal naar de basale ganglia van de emotie-organen via haar
middelste kern. En er lopen ook veel verbindingen naar de cortex.
De ui-vormige structuur rond de thalamus is
de caudate nucleus, maar die ligt al middenin de emotie-organen.
Verder met de emotie-organen hier
.
Naar Neurologie, organisatie
,
Neurologie, overzicht, globaal
,
of site home
 .
.
|
19 nov.2011; 3 mrt.2013; 2 apr.2013; 8 mei 2013; 4 sep.2014; 12 okt.2014;
15 mei 2016; 25 jun.2016; 22 jul.2016 |
|