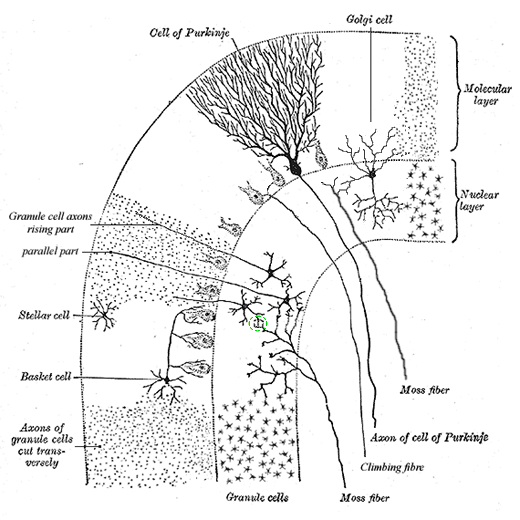
Toelichting bij Neurologie, cerebellum: schemaIn Neurologie, cerebellum is de globale informatie omtrent het neuron-netwerk in de kleine hersenen al gegeven. Hier volgt nog wat meer details. Eerst nogmaals het schetsmatige overzicht van de cerebellaire cortex:
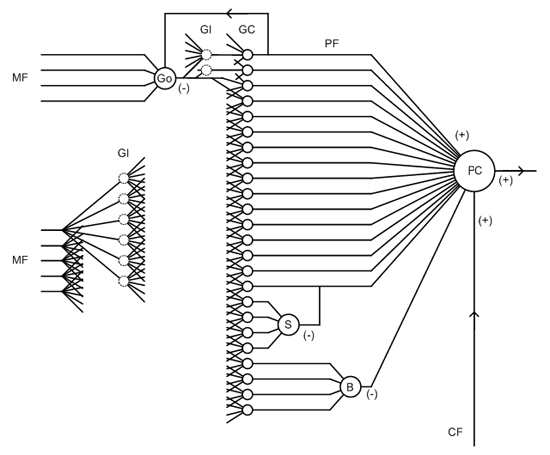
De locatie van de diverse cellen geeft ook een duidelijke aanwijzing van hun functie. De Golgi-cellen liggen in de granular of nuclear layer, tussen de granular-cellen - met hun dendrieten reikende naar de parallel fibers van de molecular layer. Dit geeft aan dat een deel van hun functie een terugkoppeling van de werking van de granular-cellen is - omdat dit ook een inhibitory effect betreft, wordt dit feedback inhibition genoemd. De eveneens remmende stellate- of stellar-cellen liggen in de molecular layer en hebben geen verbindingen naar buiten. Deze cellen oefenen hun remmende invloed dus direct op de granular-cellen - dit feed-forward inhibition. De basket-cellen liggen ook in de molecular layer, maar sturen hun output naar de Purkinje layer, de laag van Purkinje-cellichamen die tussen de molecular layer en granular of nuclear layer in ligt. Ze synapsen direct op het lichaam van de Purkinje-cel. Dit alles is gerepresenteerd in het logische schema, hier in wat een wat ruimer formaat. De exciterende of inhiberende werking van de verschillende synapsen is op de standaardwijze aangegeven met (+) en (-):
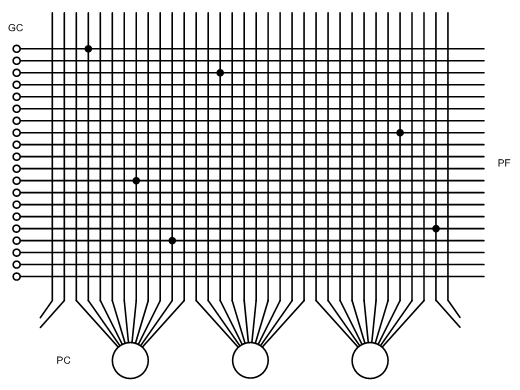
De signalen lopen van links naar rechts, tenzij anders aangegeven. De grote massa loopt van de mossy fibers via de glomeruli (naar de korrelcellen en uiteindelijk naar de Purkinje-cellen. In het schema zijn vanuit de Golgi-cel verbindingen naar zowel glomeruli als naar het cellichaam van de granular cells getekend, omdat beide in de literatuur vermeld staan - of het laatste juist is, is niet duidelijk. Ondanks de modulerende werking van de andere celsoorten zou dat een betrekkelijk simpele netwerk zijn. Gezien de rol van de taak die het cerebellum vermoedelijk verricht, ruimtelijke berekeningen en coördinatie, is een meer ingewikkelde werking voor de handliggend. Hoe dat precies gaat, is vooralsnog onduidelijk. Maar bovenstaande schema kan wel als uitgangspunt dienen voor verdere ontwikkelingen. Wel bekend is de fysieke rol van de tweede soort input naast de mossy fibres,de climbing fibres. Die hechten direct aan het lichaam van de Purkinje-cellen, en doen deze heftig vuren. Dit wordt door sommigen geassocieerd met een leerproces, maar het is onduidelijk hoe dit sterke uitgangssignaal het voorgaande deel van het netwerk beïnvloedt - zie het schema. De climbing fibre komen van de kern genaamd de inferior olivary nucleus (inferoir: onderste) in de olive, een onderdeel van het middenbrein. En technicus zou dit eerder interpreteren als een soort noodknop: op deze manier wordt de rest van het netwerk overslagen en direct naar de uitgang gegaan. Een aanlerende functie zou het actieve deel van het netwerk moeten kunnen beïnvloeden. En die toeging wordt verschaft door de mossy fibres. Het is voor de hand liggende dat een deel daarvan de aanlerende functie heeft. Daarom terug naar de basis van het netwerk. Dat bestaat uit de combinatie van granular- en Purkinje-cellen. Die combineren in de molecular layer, het dikste deel van de cortex, waarin de axonen van de granular-cellen, onder naam parallel fibers, synapsen aan de dendrieten van de Purkinje-cellen - voor één parallel fibers om ongeveer één op de vijf Purkinje-cellen. Ook dit kan het beste samengevat worden in een schema:
De algemeen geldende afspraak in dit soort schema's is dat twee lijnen die elkaar kruisen op de tekening geen verbinding maken - tenzij er een bolletje getekend is op het kruispunt. De vraag is: hoe kan in zo'n simpel netwerk, een verbindingsmatrix, een lerende functie ingebouwd zijn? Daarvoor zijn er toch meerdere mogelijkheden. Ten eerst is daar een functie die bekend is van de grote hersenen, onder de naam "use it or lose it" - dat wil zeggen: als niet-genetische bepaalde, min of meer aangeleerde, functionaliteiten, niet regelmatig gebruikt worden, gaan die functionaliteiten achteruit - dat wil zeggen: de bijbehorende neurale verbindingen worden zwakker. Dat proces kan je omkeren, en veronderstellen dat het veelvuldig gebruik van bepaalde verbindingen die verbindingen versterkt. Een tweede mogelijkheid is externe feedback. De mossy fibers komen ook van de middenbrein en hersenstam, waar de basale waarnemings- en motorische informatie rondgaat. Stel dat dat betekent dat een bepaalde dendriet van een Purkinje-cel veelvuldig geëxciteerd wordt - overeenkomstig met een bepaalde beweging. Dan is het goed voorstelbaar dat ook andere verbindingen aan deze dendriet, afkomstig van andere mossy fibers, bijvoorbeeld afkomstig uit de neo-cortex en bestaande uit: "Doe dat", versterkt worden. In die laatste verbinding is dan dus geleerd. Voor dit soort functionaliteit spelen de details van de input- en outputsignalen een cruciale rol. Die informatie staat vaak verspreid over verschillende onderwerpen - zeker in de specialistische maar ook in de algemene literatuur als Wikipedia. Hier een poging tot verzameling. Voor die verzameling stappen we weer over op de evolutionaire benadering. Het cerebellum is namelijk een culminatie van eerdere ontwikkelingen, die plaats hebben gevonden vanaf het ruggemerg. Daar gaan we dus beginnen. Het neurologisch ruggemerg is iets dat stamt van alle gewervelde dieren. Het is een verzamelplaats voor zenuwbanen komende van waarnemingsorganen, en gaande naar spieren van ledematen en elders. In het ruggemerg vinden de eerste besturingsprocessen plaats:de coördinatie van de onderdelen van de ledematen, en de combinatie met de input van de waarnemingsorganen. De coördinatie van onderdelen van ledematen is al een punt bij zoiets simpels als een tentakel - om die als geheel te kunnen sturen, moeten er vele spieren op een gecoördineerde manier samenwerken. De door de mens gebouwde machines komen op dat terrein nog niet verder dan wat bijzonder houterig uitziend gestuntel. Die coördinatie vindt plaats in zenuwknopen in het ruggemerg, in klompjes neuronen genaamd ganglia. Bij de coördinatie van spieren komt nog een tweede essentieel verschijnsel om de hoek kijken: terugkoppeling. Het tweede is de coördinatie tussen waarnemingsorganen en besturing. Dat is ingewikkelder, vergt meer neuronen, en meer plaats dan er is in het ruggemerg van een gewervelde. De eerste vormen van coördinatie zijn van de soort: zie zwarte vlek van links naar rechts op het netvlies, steek tong uit (in mensentaal: "kikker vangt vlieg"). Nu is zelfs deze basale handeling al bijzonder ingewikkeld, want de tong moet niet uitgestoken worden richting zwarte vlek, maar ergens daar waar de zwarte vlek zal zijn als de tong eenmaal volledig is uitgerold. Dat is dusdanig ingewikkeld, en de knoop van neuronen die dit coördineert dusdanig groot, dat deze al snel buiten het ruggemerg komt te liggen - dat is de eerste vorm van het brein. Zo'n grote knoop met een duidelijk samenhangende structuur heet een nucleus. Diverse nuclei met bijbehorende functies vormen neurologische organen. En de verzameling van deze organen is het brein. Waarbij er in het brein diverse groeistadia zijn voorgekomen, leidende tot de driedeling: hersenstam, emotionele hersenen, grote hersenen. Hier gaat het dus alleen om de fase van ruggemerg naar hersenstam, maar dat is de fase die volledig zelfstandig het lichaam kan besturen. Wat daar boven gebeurt is slechts een aanpassing op dat volledige functioneren - het verschuift gedragingen, maar maakt relatief slechts een zeer beperkt aantal nieuwe functionaliteiten aan. Ten opzichte van het menselijke functioneren als geheel is er weinig verschil tussen het met een groepje van tien een boom uithollen en met een groepje van honderd een VOC-schip bouwen. Naar Neurologie, cerebellum
|